microRNA甲基化修饰
m6A是真核生物中较常见的RNA修饰,被认为是一种新的表观遗传标记,参与多种生物过程。m6A调控几种主要人类病毒性疾病的模式和功能已经有报道。然而,m6A在植物病害暴发中的分布模式和作用尚不清楚。在此,作者分析了两种破坏性的病毒感ran水稻植株中的m6A修饰情况。发现m6A甲基化主要与病毒gan染水稻植株中表达不活跃的基因有关。作者还检测到同一基因上不同的m6A峰分布,这可能是水稻条纹病毒和水稻黑条纹矮病毒感ran之间存在不同抗病毒模式的原因。有趣的是,病毒感ran后水稻m6A甲基化水平增加。环状RNA也会被m6A甲基化修饰,并会促进环状RNA编码蛋白这一有趣的结论。microRNA甲基化修饰
植物m6A表观转录组修饰的生物学功能尚不完全清楚。CPSF30-L是聚腺苷酸化因子CPSF30的主要亚型,由CPSF30-S和一个m6A结合的YTH结构域组成。CPSF30-L的生物学作用及其在选择性聚腺苷酸化中结合m6A功能的分子机制尚不清楚。在这里,作者揭示了CPSF30-L是拟南芥m6A的“reader”,其m6A结合功能是花卉的转型和脱落酸(ABA)响应所必需的。此外,CPSF30-L的m6A结合活性增强了液体样核体的形成,其中CPSF30-L主要识别m6A修饰的远上游元件,调控聚腺苷酸化位点的选择。mRNA甲基化利器m6A除了分布在 mRNA 中,也出现在很多非编码 RNA中 ,如:环状RNA 、LncRNA等。
m6A修饰在植物中是必不可少的。在这里,作者证明了在水稻中表达转基因人RNA去甲基化酶FTO可以使温室条件下的粮食产量增加3倍以上。在田间试验中,转基因FTO在水稻和马铃薯中的表达使产量和生物量增加了约50%。此外,FTO的存在促进了根分生组织细胞增殖和分蘖芽的形成,提高了光合效率和抗旱性,但对成熟细胞大小、茎分生组织细胞增殖、根直径、株高或倍性没有影响。FTO介导了植物RNA中大量的m6A去甲基化(在poly(A) RNA中约7%的去甲基化,在非核糖体核RNA中m6A下降约35%),诱导染色质开放和转录激huo。因此,调控植物RNA m6A甲基化是一种很有前景的策略,可明显提高植物的生长和作物产量。
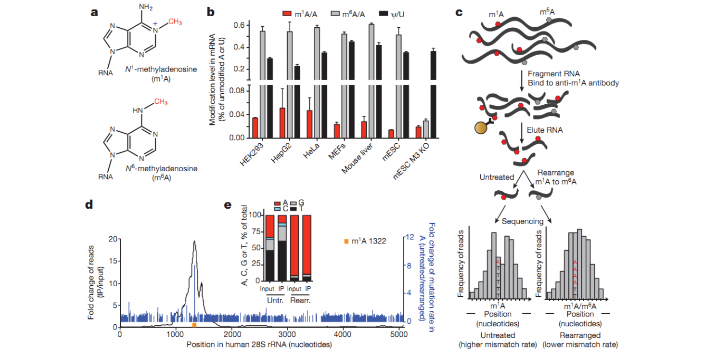
RNA m6A甲基化是RNA较关键的内部修饰之一,是真核生物丰富和调节遗传信息的一种保守的转录后机制。m6A修饰具有多种生物学功能,如调控mRNA翻译、转录以及稳定性等。目前,已在多种植物中发现这种RNA修饰,参与调控不同植物的各方面生物学功能。其中,拟南芥作为植物界中研究RNA甲基化修饰的先行者,许多学者将它作为研究对象,并与m6A甲基化测序技术结合,证实到RNA甲基化普遍存在于拟南芥各个发育期,并揭示了RNA甲基化相关酶在特殊发育时期,如开花,叶片形成,种子发育,根部生长等过程中发挥重要作用。RNA m6A是真核生物丰富和调节遗传信息的一种保守的转录后机制。
m7G RNA甲基化是在甲基化转移酶的作用下,使RNA鸟嘌呤(G)的第七位N上加上甲基的一种修饰( N7-methylguanosine,m7G)。研究表明,m7G RNA甲基化修饰存在于各类分子中,包括:mRNA 5’帽子结构、mRNA内部、pri-miRNA、转运RNA(tRNA)和核糖体RNA(rRNA)。m7G RNA甲基化修饰能够调节mRNA的转录、miRNA的生物合成和生物学功能、tRNA稳定性、18S rRNA的核内加工及成熟。m7G RNA甲基化修饰作为一类新型RNA甲基化,近两年高影响因子文章不断,是继m6A修饰之后的又一表观转录组学热点。RNA去甲基化可以提高水稻和马铃薯植株的产量和生物量。浙江tRNA甲基化
比色法RNA甲基化定量检测试剂盒提供了定量检测总RNA甲基化水平的试剂。microRNA甲基化修饰
ac4C RNA乙酰化是在RNA ac4C修饰酶的作用下,使N4位乙酰胞嘧啶发生乙酰化的一种保守的化学修饰(N4-acetylcytidine)。早期研究发现该修饰存在于真核生物中丝氨酸及亮氨酸tRNA和18S rRNA上,导致Watson-Crick碱基热稳定性增加,调控蛋白合成中的编码准确性;近期研究发现ac4C分布在人类转录组中,大多数位点出现在编码序列(CDS)内,并且通过改善的mRNA稳定性和翻译促进靶基因表达。目前,ac4C RNA乙酰化修饰作为一类新型RNA修饰,是继m6A修饰之后的又一表观转录组学热点和“超级潜力股”!microRNA甲基化修饰
上一篇: dna的甲基化的检测方法
下一篇: 广西miRNA甲基化